Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2013/2018
[A propos de l'auteur]
[Droits de copie]
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2013/2018
[A propos de l'auteur]
[Droits de copie]
On va modéliser le chémostat à l'aide d'une série d'équations différentielles simples. Ce système a été posé dans un article historique de Monod en 1950 (J. Monod, La technique de la culture continue: Théorie et applications, Ann. Inst. Pasteur, Lille, 79 (1950), 390). Dans ce modèle simple fondamental, les cellules se multiplient et on néglige toute mortalité cellulaire et on n'envisage aucune production d'un produit quelconque dans le milieu par les cellules. De même, aucun inhibiteur n'apparaît.
On suppose un chemostat dans lequel un des nutriments appelons le SL va être le nutriment qui va limiter la valeur de la vitesse spécifique de croissance (µ) de la biomasse.
Les autres nutriments étant à des concentrations suffisament élevées pour ne pas exercer ce même effet de limitation. L'équation qui règle la limitation de µ est :

Regardons ce qui se passe alors pour la biomasse ([X]) dans le bioréacteur :

On va ainsi écrire :
![]() Ce sera l'équation 2
Ce sera l'équation 2
Regardons finalement le bilan pour les différents substrats S (valable aussi pour Sl, le substrat limitant de µ) :

On va écrire cette équation sous la forme qui suit en remplaçant rX par µ[X] puique rX = µ[X]:
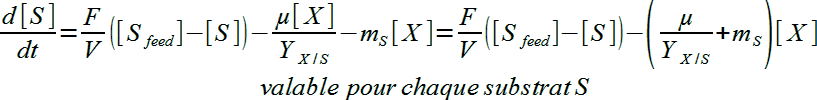 Ce sera l'équation 3
Ce sera l'équation 3
L'équation (3) est évidemment valable pour le substrat SL qui limite µ ! Et on l'appliquera dans l'exemple pratique du $3. On peut aussi mentionner que mS, coefficient de maintenance est souvent (pas toujours !) très faible devant µ/YX/S et donc, le terme lié à la maintenance peut souvent être négligé dans l'équation (3).
Regroupons nos 3 équations pour bien visualiser le modèle de gouvernance du chémostat :
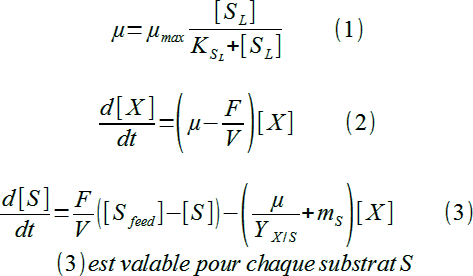
Un système bien "équilibré" puisqu'on a par exemple 3 inconnus : µ, X et Sl (substrat limitant de µ) pour 3 équations ! Les autres éléments sont, en effet, des paramètres physiologiques de la souche en culture (µmax, K0,5Sl, YX/Sl, mSl) ou des paramètres de "construction" du chémostat ([Sl-feed], F et V).
La suite, pratique, au paragraphe 3...