Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2012-2020
[A propos de l'auteur]
[Droits de copie]
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2012-2020
[A propos de l'auteur]
[Droits de copie]
Globalement, dans une fermentation classique, le nutriment énergétique (le plus souvent un bon vieux sucre) propose de quoi être oxydé contre des coenzymes transporteurs d'électrons qui vont ainsi passer de l'état oxydé à réduit. Ces redox à l'enthalpie libre très favorable (très négative) fournissent l'énergie qui permet de générer les fameux composés à haut potentiel d'hydrolyse (anhydrides d'acide phosphorique et autres énols phosphate). L'ATP sera alors régénéré par (ADP)phosphorylation au niveau du substrat. Gros problème : la cellule se retrouve rapidement avec un stock de coenzymes transporteurs d'électrons à l'état réduit (NADH en général) sur les bras. Il va bien falloir les réoxyder pour pouvoir fermenter de nouveau substrats. Ce recyclage des coenzymes réduits sera réalisé à l'aide de ce qu'est devenu le substrat énergétique lui même pendant le processus de fermentation. Ce recyclage conduit à ce qu'on appelle les produits terminaux de fermentation (alcools divers, acides organiques divers, dioxyde de carbone, dihydrogène ...). On voit bien que pour être ainsi fermenté un substrat doit proposer de quoi être oxydé (c'est ce qui donnera l'énergie) et de quoi être réduit (c'est ce qui permettra de recycler les coenzymes réduits) : pas si simple. Et on voit ici la grande qualité des sucres pour être fermentés : ils présentent un bel équilibre de fonctions aldéhydes/cétones et alcool, pile ce qu'il fallait.
On propose le schéma de principe suivant :

On trouvera la voie dans de nombreuses pages internet, mais en voici un schéma :

• On voit alors que la source d'énergie "raffinée" est le glyceraldéhyde-3-phosphate. Le raffinage coûte d'ailleurs assez cher en énergie.
• La réaction catalysée par la glyceraldéhyde-3-phosphate deshydrogénase est remarquable. C'est l'enthalpie libre très négative de la réaction d'oxydation du glyceraldéhyde-3-phosphate au niveau de sa fonction aldéhyde qui va être couplée à l'incorporation d'un phosphate et à la création de l'anhydride d'acide phosphorique 1,3-bisphosphoglycérate. C'est ainsi qu'apparaît le 1,3-bisphosphoglycérate molécule pleine d'énergie utilisable pour réaliser des phosphorylations au niveau du substrat. La réaction conduit à du NADH (du NAD réduit).
• Une première réaction de phosphorylation au niveau du substrat peut intervenir directement sur le 1,3-bisphosphoglycérate. Une deuxième sur le phosphoénolpyruvate qui découle ensuite. Les deux enzymes qui catalysent ces 2 réactions sont la 3-phosphoglycérate kinase et la pyruvate kinase. On obtient finalement du pyruvate. En considérant le glucose de départ, le bilan est positif de 2 ATP par glucose et on obtient 2 pyruvate.
• Comment recycler le NADH ? Chez les bactéries lactiques homofermentaires, tout simplement par réduction du pyruvate en lactate selon :
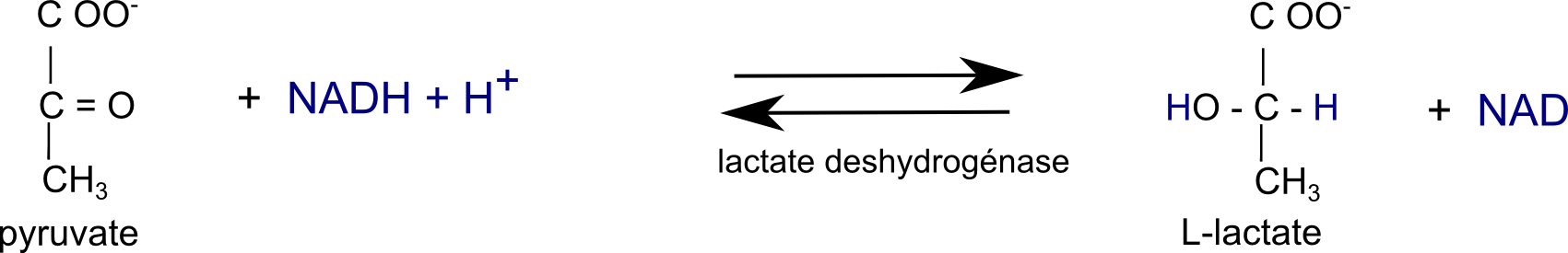
Ainsi chez les bactéries lactiques homofermentaires, il se forme environ 1,8 mole de lactate par mole de glucose consommé. Pas tout à fait 2 évidemment (1 glucose donne 2 pyruvates) puisqu'une partie du pyruvate peut être recyclé comme source de carbone
, utilisation qui nécessite d'ailleurs des réductions qui permettront "d'éponger" des coenzymes réduits (pratique !).
• C'est la célèbre fermentation alcoolique des levures.
• L'analyse est à l'identique de celle ci-dessus sauf pour le recyclage du NADH qui utilise la fameuse séquence pyruvate décarboxylase / éthanol deshydrogénase :

Le schéma ci-dessous présente la voie hétérolactique (lactique hétérofermentaire) de fermentation du glucose. On trouvera aussi la description de cette voie dans de nombreuses pages internet, par exemple au sein de ce document assez complet du "Todar's online textbook of mmicrobiology" traitant des fermentations : ouvrir un nouvel onglet à http://textbookofbacteriology.net/metabolism_3.html.

• Par rapport à la voie EMP, cette voie se caractérise par l'absence de fructose-1,6-biphosphate aldolase (qui scinde le fructose-1,6-biphosphate en dihydroxyacétone-phosphate et glyceraldéhyde-3-phosphate) et par la présence de l'activité phosphocétolase sur le ribulose-5-phosphate qui produit du glyceraldéhyde-3-phosphate et de l'acétyl-phosphate, 2 composés à haut potentiel d'hydrolyse ! Faisant suite à 2 oxydations avec réduction de NAD+ en NADH, c'est elle qui est à la source des composés à haut potentiel de cette fermentation selon une réaction phosphorylante remarquable.
• Comment recycler le NADH ? Chez les bactéries lactiques hétérofermentaires ? Par la très classique lactate déshydrogénase qui réduit le pyruvate en lactate mais aussi par la série de réactions :
acétyl-phosphate + coA-SH ---> acétyl-CoA + phosphate
puis acétyl-CoA + NADH + H+ ---> acétaldéhyde + NAD+
puis acétaldéhyde + NADH + H+ ---> éthanol + NAD+
Globalement on réoxyde 2 NADH moyennant la perte d'un composé à haut potentiel d'hydrolyse (l'acétyl-phosphate).
• Classiquement, pour équilibrer la voie entre oxydation et réduction au niveau NAD+/NADH, il est usuel de schématiser la voie comme si tout l'acétyl-phosphate allait vers l'éthanol. Mais les choses sont un peu plus complexes que ça dans la nature puisque des réactions de biosynthèse consomment du pouvoir réducteur et qu'ainsi une fraction non négligeable de l'acétyl-phosphate peut conduire à de l'acétate par phosphorylation au niveau du substrat avec un ATP de gagné. Les principaux produits terminaux de fermentation sont donc : le CO2, le lactate, l'éthanol et l'acétate avec une prédominance du lactate sur l'éthanol et l'acétate.