Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2004-2021
[A propos de l'auteur]
[Droits de copie]
![]()
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour 2004-2021
[A propos de l'auteur]
[Droits de copie]
![]()

DH2 = nutriment énergétique catabolisé = donneur d'électrons. Une série de réactions enzymatiques vont conduire à son oxydation contre des Coenzymes transporteurs d'électrons qui vont passer de l'état oxydé (T) à l'état réduit (T2H+2e).

D = déchet du métabolisme résultant de l'oxydation du nutriment énergétique
DH2.
Exemple : DH2 = glucose et D = CO2 (par voie EMP puis cycle de Krebs)
chez de nombreuses bactéries chmiorganotrophes à respiration aérobie.
Phase 1 L'oxydation du donneur d'électrons DH2 est couplée à la réduction de coenzymes transporteurs d'électrons qui passent ainsi de l'état oxydé T à l'état réduit T2H+2e.
La phase 1 a été placée en phase "soluble" sur le schéma car elle est essentiellement en phase "soluble" lors des respirations les plus classiques à accepteur O2 ou NO3-. Mais les réactions à enzymes membranaires sont très classiques : pour n'en citer qu'une à ce niveau de l'exposé, la succinate déshydrogénase ...
Un exemple type : dans les respirations aérobies classiques sur glucose, le glucose est oxydé en CO2.

T = Coenzyme transporteur d'électrons à l'état oxydé.
Les coenzymes transporteurs d'électrons sont classiquement le couple NAD+/NADH,H+ (passage sous forme libre soluble) et FAD/FADH2 (groupement prosthétique d'enzymes membranaires).

T2H+2e = Coenzyme transporteur d'électrons à l'état réduit.
Les coenzymes transporteurs d'électrons sont classiquement le couple NAD+/NADH,H+ (passage sous forme libre soluble) et FAD/FADH2 (groupement prosthétique d'enzymes membranaires).
Phase 2 en phase membranaire.
Les coenzymes réduits vont être réoxydés au niveau des complexes protéiques
d'une chaîne respiratoire. Le bilan redox est :
T2H+2e + A ---> T + AH2
ΔG<<0
L'enthalpie libre n'est pas perdue en chaleur, elle est convertie en
translocation de protons à travers la membrane, c'est à dire convertie en
potentiel électrochimique : le gradient protonmoteur. Dans les ouvrages on parle de couplage
chimio-osmotique (un terme historique pas heureux du tout) ou de couplage chimio-électrochimique.
Les chaînes respiratoires (CR) sont formées de complexes protéiques membranaires ; on y trouve des protéines de nature cytochromique.
Chez les bactéries à métabolisme respiratoire, la membrane lieu des CR est évidemment la membrane cellulaire. Chez les eucaryotes, il s'agit de la membrane interne des mitochondries
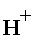
LeΔG<<0 de certaines des réactions redox de la chaîne respiratoire n'est pas perdu en chaleur mais est couplé à la translocation de protons = couplage chimio-électrochimique. = mise en place d'un gradient protonmoteur transmembranaire.
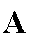
A = Accepteur terminal des électrons. L'accepteur terminal
des électrons de la chaîne respiratoire est réduit côté intracellulaire chez les procaryotes dans les cas
les plus classiques (O2 ou NO3-
comme accepteur) et côté matrice mitochondriale chez les eucaryotes (O2 est l'unique accepteur chez les eucaryotes). Mais
les microorganismes sont très inventifs ...
Le plus classique des accepteurs est O2 chez les aérobies :
O2 + 4e en provenance de la chaîne respiratoire + 4 H+ donne 2 H2O.
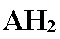
AH2 : A (accepteur terminal des électrons) a été réduit en AH2.
Le bilan redox des phases 1 et 2 est :
DH2 + A ---> D + AH2
ΔG<<0
L'enthalpie libre n'est pas totalement perdue en chaleur, une partie est convertie, au niveau de la CR en
translocation de protons à travers la membrane, c'est à dire convertie en
potentiel électrochimique : le gradient protonmoteur.
Dans le cadre du schéma proposé O2 est l'accepteur terminal des électrons :
O2 + 4e en provenance de la chaîne respiratoire + 4 H+ donne 2 H2O.
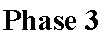
Phase 3. Le potentiel électro-chimique membranaire obtenu par la translocation des protons en phase 2 va être couplé à la régénération d'ATP grâce aux ATPsynthases membranaires. La membrane est imperméable aux protons. Les ATPsynthases vont permettre le passage des protons dans le sens de leur gradient électro-chimique avec utilisation du ΔG très négatif associé pour générer une liaison anhydride d'acide phosphorique entre ADP et Pi. C'est un couplage électro-chimique chimique.
Le mécanisme global Phase2 + Phase3 est appelé phosphorylation oxydative.
Note importante : la réaction est parfaitement réversible.
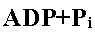


Les ATPsynthases (ou ATPases) membranaires permettent l'interconcversion "monnaie énergétique ATP" et "monnaie énergétique gradient protonmoteur".

Mmbrne désigne ici la membrane cellulaire chez les procaryotes, la membrane interne mitochondriale chez les eucaryotes. Elle est étanche aux protons.
Intérieur signifie ici : cytoplasme cellulaire chez les procaryotes, matrice mitochondriale chez les eucaryotes.
Extérieur signifie ici : espace périplasmique chez les procaryotes, espace inter-membranaire mitochondrial chez les eucaryotes.

Intérieur signifie globalement que ça se passe "classiquement" côté cytoplasme cellulaire. Mais les micro-organismes sont inventifs ... (et essentiellement dans la matrice mitochondriale chez les eucaryotes !)
Mmbrne désigne ici la membrane cellulaire chez les procaryotes, la membrane interne mitochondriale chez les eucaryotes. Elle est étanche aux protons.
Extérieur signifie ici : espace périplasmique chez les procaryotes, espace inter-membranaire mitochondrial chez les eucaryotes.

Extérieur signifie ici : espace périplasmique chez les procaryotes, espace intermembranaire mitochondrial chez les eucaryotes.
Intérieur signifie ici : cytoplasme cellulaire chez les procaryotes, matrice mitochondriale chez les eucaryotes.
Mmbrne désigne ici la membrane cellulaire chez les procaryotes, la membrane interne mitochondriale chez les eucaryotes. Elle est étanche aux protons.
• Si ce schéma de respiration s'applique généralement bien aux respirations aérobies avec source primaire organique. O2 est ainsi l'accepteur
terminal des électrons. Parfois, en anaérobiose, ce schéma n'est pas de portée universelle !! (Se reporter à la page définissant le terme "métabolisme énergétique respiratoire"
respirations_1.php et aux exemples particuliers
présentés dans la suite.)
• Contre exemple 1. Les bactéries acétiques qui utilisent la source primaire d'énergie éthanol et rejettent de l'acide acétique le font par respiration aérobie mais
l'éthanol est directement donneur d'électrons à un complexe membranaire cytochromique respiratoire sur la face externe de la membrane cellulaire... Voir
des détails au paragraphe 6 ; http://perrin33.com/microbiologie/metabolismes/respirations_6.php.
• Autre contre exemple. Si on s'intéresse de près aux métabolismes énergétiques généralement qualifiés de respirations sulfate (SO42- accepteur
terminal), les phases 1 et 2 devront être un peu revues...
• Et encore. Le métabolisme aérobie sur substrat pyrite de bactéries comme Thiobacillus ferrooxydans est bien considéré
comme respiratoire avec fe2+ (nutriment énergétique) donneur direct d'électrons à la chaîne respiratoire par le côté périplasmique ! ... voir
des détails au paragraphe 7 ;
http://www.perrin33.com/microbiologie/metabolismes/respirations_7.php.
• Et pour finir. Le métabolisme énergétique des méthanogènes présente
des aspects qui feront penser aux respirations mais l'origine du gradient proton-moteur est tout de même très particulière ... Personnellement
le métabolisme méthanogène mériterait pour moi une appellation en soi (ni fermentation, ni respiration). Certains auteurs classent la méthanogénèse dans les respirations (ce qui me
paraît être un peu olé olé...) et il est souvent d'usage de dire "fermentation méthanogène" puisque, par convention, tout ce qui n'est pas phototrophie et n'est pas
respiration est qualifié de fermentation.
Il faut rester ouvert à la diversité du vivant et ne pas faire de ce schéma un dogme.