Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour octobre 2015
[A propos de l'auteur]
[Droits de copie]
![]()
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour octobre 2015
[A propos de l'auteur]
[Droits de copie]
![]()
On suppose un bioréacteur qui produit de la biomasse (X). On suppose un métabolisme chimiotrophe. On s'intéresse à la consommation d'un substrat S.
A priori, on peut supposer que du S est consommé sur le poste de la maintenance cellulaire (le fait de vivre consomme du S) et sur le poste croissance (pour former de la biomasse nouvelle, il faut consommer du S). On peut donc imaginer un modèle de consommation très simple somme de ces 2 postes.
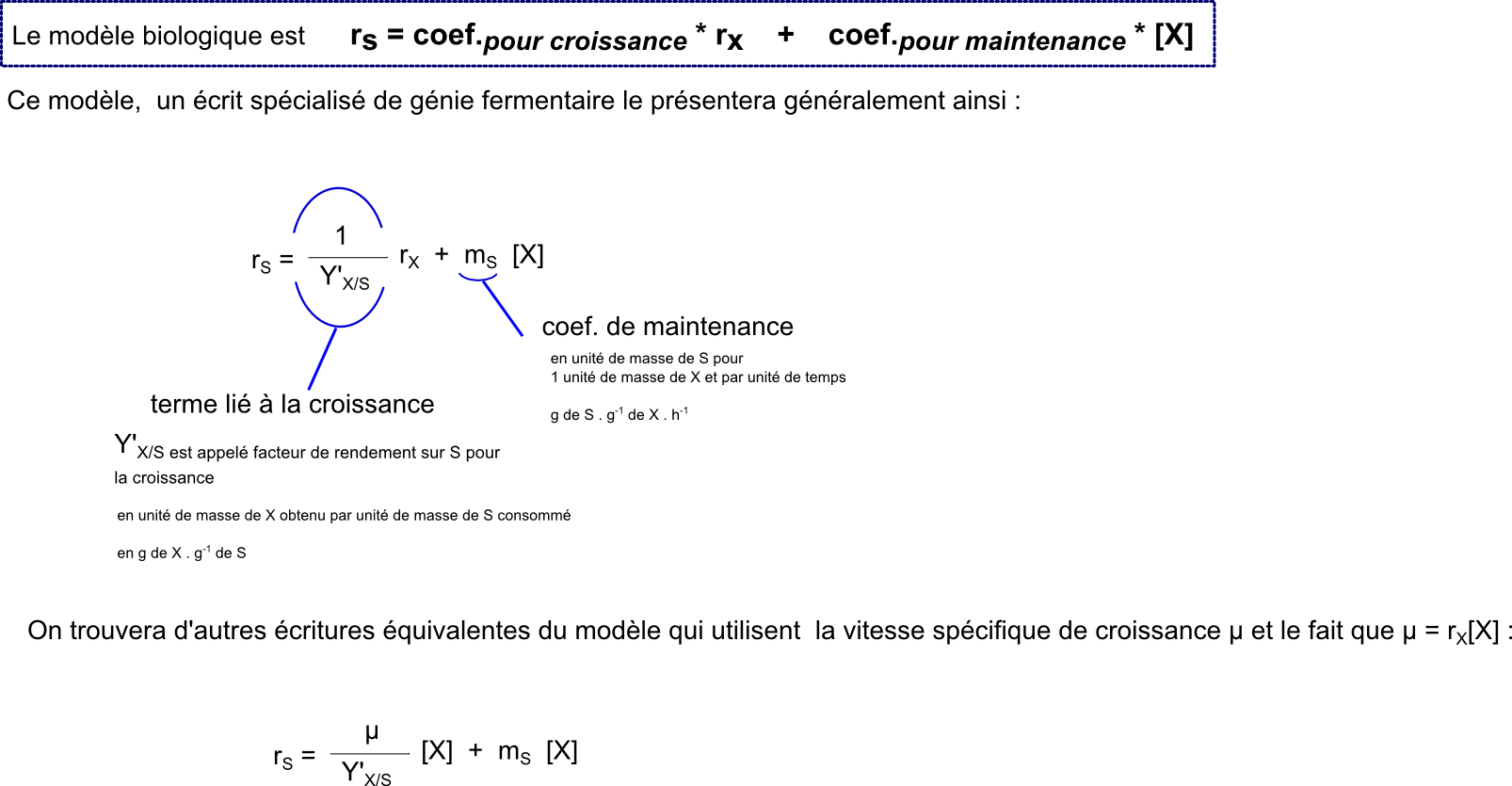
Et une remarque fondamentale, le rendement de conversion de S en X ne pourra évidemment jamais être meilleur que le rendement donné par la relation stoechiométrique de conversion de S en X.
Expliquons cette affaire sur 2 exemples à partir du cas d'un bioréacteur qui produit de la biomasse (X) avec un métabolisme chimioorganotrophe sur substrat glucose avec une unique source d'azote nitrate.
Exemple 1.
Supposons une composition cellulaire élémentaire déterminée et - rapportée à 1 C - égale à CH1,9O0,4N0,28P0,02 plus 4% de la masse sous forme de minéraux.
On en déduit une masse atomique relative de (12+1,9x1+0,4x16+0,28x14+0,02x31)x100/96=26 pour chaque élément carbone de la biomasse.
Ainsi, selon les données de composition élémentaire de la biomasse 1 glucose (C6H12O6,
soit 180 de masse moléculaire relative et 6 atomes de C) a un "potentiel stoechiométrique" de 6*26 = 156 g de biomasse.
Ce "potentiel stoechiométrique" correspond ainsi à un rendement stoechiométrique de conversion de S en X de
156/180 = 0,87 de biomasse g-1 de glucose.
La valeur 0,87 est le résultat d'un calcul stoechiométrique limite qui fait comme si tout le C du glucose se retrouvait en biomasse et qui ne conçoit donc le glucose que comme source de C et qui supposerait l'intégration du C du glucose en biomasse sans aucune perte. Ceci est évidemment inexact in vivo puisque le glucose est notamment source d'énergie. Il y aura donc un écart entre le rendement stoechiométrique calculé et le rendement réel biologique qu'on pourra mesurer. Par exemple, on mesurera un rendement réel vers 0,5 g de biomasse g-1 de glucose (voir §2).
On pourra en déduire que 0,5/0,87 = 57% du glucose consommé lors la croissance l'est comme source de C (C retrouvé dans la biomasse créée) alors que 43 % n'est pas retrouvé dans la biomasse mais a essentiellement servi de source d'énergie.
Exemple 2.
Reprenons la composition cellulaire élémentaire précédente : CH1,9O0,4N0,28P0,02 plus 4% de la masse sous forme de minéraux. Soit en se ramenant à 1N : C3,6H6,8O1,5NP0,072 plus 4% de la masse sous forme de minéraux.
On en déduit une masse atomique relative de (3,6x12 +6,8x1+1,5x16+14+0,072x31)x100/96= 94 pour chaque élément azote de la biomasse.
Ainsi, selon les données de composition élémentaire de la biomasse 1 NO3- (
62 de masse moléculaire relative pour 1 N) a un "potentiel stoechiométrique" de 94 g de biomasse.
Ce "potentiel stoechiométrique" correspond ainsi à un rendement stoechiométrique de de conversion de NO3- en X
de 94/62 = 1,5 de biomasse g-1 de NO3-.
La valeur 1,5 est le résultat d'un calcul stoechiométrique qui suppose que tout l'azote de chaque nitrate se retrouve en biomasse et qui ne conçoit donc le nitrate que comme source d'azote et qui suppose l'intégration de l'azote en biomasse sans aucune perte. Ceci sera a prioriinexact in vivo puisque même si le nitrate n'est que source d'azote, on peut supposer des pertes liés au turn-over des molécules. Il y aura donc un écart entre le rendement stoechiométrique calculé et le rendement réel. Mais on verra au §3 que la consommation de NO3- pour la maintenance est quasi négligeable devant celle pour la croissance lors d'une culture classique. Le rendement stoechiométrique sera quasiment atteint au cours de la croissance.