Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour octobre 2015
[A propos de l'auteur]
[Droits de copie]
![]()
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour octobre 2015
[A propos de l'auteur]
[Droits de copie]
![]()
Soit par exemple une culture discontinue (en batch) ayant donné les mesures proposées par les 3 colonnes "bleues" puis les valeurs calculées approchés proposées par les colonnes "jaunes" :
Temps t (h) |
biomasse X |
[glucose] |
Calcul approché de |
Calcul approché de |
D'où la détermination de |
D'où la détermination de |
0 | 0.100 | 40 |
|
| ||
1 | 0.134 | 39.93 | (0,180-0,100)/2=0,040 |
(40,00-39,83)/2=0,085 | =0,040/0,134=0,298 | =0,085/0,134=0,634 |
2 | 0,180 | 39,83 | 0,054 |
0,115 | 0,300 | 0,639 |
3 | 0,241 | 39,70 | 0,072 |
0,165 | 0,299 | 0,687 |
4 | 0,373 | 39,50 | 0,096 |
0,200 | 0,297 | 0,619 |
5 | 0,433 | 39,30 | 0,129 |
0,265 | 0,298 | 0,612 |
6 | 0,581 | 38,97 | 0,172 |
0,400 | 0,296 | 0,688 |
7 | 0,778 | 38,50 | 0,230 |
0,485 | 0,296 | 0,623 |
8 | 1,040 | 38,00 | 0,311 |
0,650 | 0,299 | 0,625 |
9 | 1,400 | 37,20 | 0,415 |
0,900 | 0,296 | 0,643 |
10 | 1,870 | 36,20 | 0,550 |
1,200 | 0,294 | 0,642 |
11 | 2,500 | 34,80 | 0,740 |
1,650 | 0,296 | 0,660 |
12 | 3,350 | 32,90 | 0,995 |
2,150 | 0,297 | 0,642 |
13 | 4,490 | 30,50 | 1,325 |
2,850 | 0,295 | 0,635 |
14 | 6,00 | 27,20 | 1,755 |
3,850 | 0,293 | 0,642 |
15 | 8,00 | 22,80 | 2,350 |
5,050 | 0,294 | 0,631 |
16 | 10,70 | 17,10 | 3,050 |
6,600 | 0,285 | 0,617 |
17 | 14,1 | 9,60 | 3,600 |
7,995 | 0,255 | 0,567 |
18 | 17,9 | 1,11 | 2,100 |
4,800 | 0,117 | 0,268 |
19 | 18,3 | 0,00 | 0,200 |
1,055 | 0,011 | 0,058 |
20 | 18,3 | 0,00 |
|
| le "x" du y=ax+b | le "y" du y=ax+b |
le tableau ci-dessus a été obtenu en adaptant partiellement la présentation de données figurant dans le chapitre fermentation modelling de Sinclair C. G., Cantero D. 1990. In: Fermentation a practical approach. Eds.: McNeil B., Harvey L. M.; IRL PRESS at Oxford University Press, New York, pp. 65-112.
On suppose que la composition élémentaire de la biomasse est CH1,9O0,4N0,28P0,02 plus 4% de la masse sous forme de minéraux.. On a vu alors au paragraphe 1 que le rendement stoechiométrique de conversion du glucose en biomasse est de 0,87 g de biomasse g-1 de glucose. Le glucose n'étant pas que source de C mais aussi source d'énergie les rendements pratiques seront donc très inférieurs à cette valeur 0,87.
On peut illustrer avec le rendement global de conversion du glucose en biomasse sur la durée de 19 h de culture qui conduit à son épuisement, YX/S global = (18,3-0,1)/(40-0) = 0,46 g de biomasse g-1 de glucose. Evidemment bien loin de 0,87.
On reprend le modèle de base exposé au paragraphe 1.
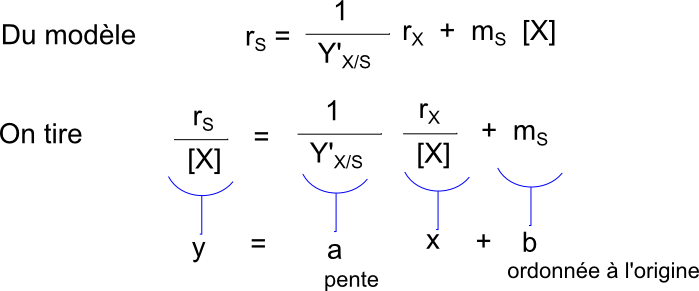
Si Y'X/S et mS sont constants en cours de culture (ce qui ne paraît pas être une hypothèse physiologiquement ridicule) on devrait obtenir une droite en testant le graphe rS/[X]=f(rX/[X]). On utilise les données du tableau du § 2.1 et on obtient :
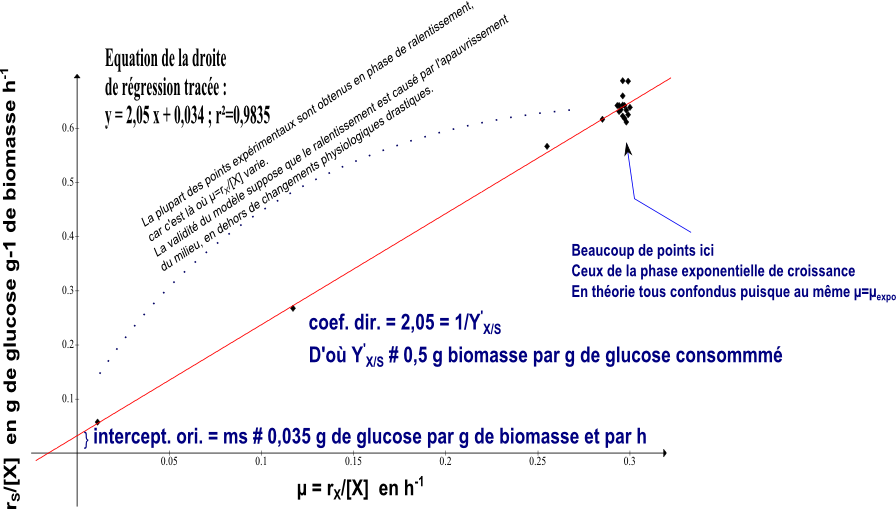
Il s'agit de bien comprendre la signification des résultats obtenus. mS=0,035 g glucose g-1 biomasse h-1 signifie que la maintenance d'un g de biomasse consomme 0,035 g de glucose par h. Le glucose étant dans l'exemple proposé source de C et d'énergie, il faut bien comprendre que ce terme de maintenance englobe 2 postes, l'énergie pour la maintenance et la consommation de C pour la maintenance, c'est à dire pour le renouvellement physiologique de la matière vivante en dehors de toute croissance. Y'X/S = 0,5 g de biomasse g-1 de glucose signifie qu'avec 1 g de glucose, les cellules peuvent créer 0,5 g de biomasse "neuve". La aussi le glucose est utilisé comme source de C et d'énergie.
On peut aussi reprendre ici la réflexion vue au paragraphe 1 concernant la comparaison rendement mesuré in vivo et rendement stoechiométrique. la voici :
De la composition cellulaire élémentaire déterminée égale à CH1,9O0,4N0,28P0,02 plus 4% de la masse sous forme de minéraux, on avait déduit que ce "potentiel stoechiométrique" correspondait à un rendement stoechiométrique de conversion de S en X de de 156/180 = 0,87 de biomasse g-1 de glucose.
La valeur 0,87 est le résultat d'un calcul stoechiométrique limite qui fait comme si tout le C du glucose se retrouvait en biomasse et qui ne conçoit donc le glucose que comme source de C et qui suppose l'intégration du C du glucose en biomasse sans aucune perte. Ceci est évidemment inexact in vivo puisque le glucose est notamment source d'énergie. Il y a donc un écart entre le rendement stoechiométrique calculé et le rendement réel biologique qu'on a pu mesurer. On a mesuré un rendement réel pour la croissance Y'X/S = 0,5 g de biomasse g-1 de glucose pendant la croissance.
On en déduit que 0,5/0,87 = 57% du glucose consommé lors la croissance l'est comme source de C (C retrouvé dans la biomasse créée) alors que 43 % n'est pas retrouvé dans la biomasse mais a essentiellement servi de source d'énergie.