Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour octobre 2015
[A propos de l'auteur]
[Droits de copie]
![]()
Retours vers :
[Accueil]
[Sommaire du dossier]
JF Perrin mise à jour octobre 2015
[A propos de l'auteur]
[Droits de copie]
![]()
Soit par exemple une culture discontinue (en batch) ayant donné les mesures proposées par les 3 colonnes "bleues" puis les valeurs calculées approchés proposées par les colonnes "jaunes" :
Temps t (h) |
biomasse X |
[NO3-] |
Calcul approché de |
Calcul approché de |
D'où la détermination de |
D'où la détermination de |
0 | 0.100 | 17,7 |
|
| ||
1 | 0.134 | 17,8 | (0,180-0,100)/2=0,040 |
non calculé |
=0,040/0,134=0,298 | non calculé |
2 | 0,180 | 17,7 | 0,054 |
0,300 | ||
3 | 0,241 | 17,6 | 0,072 |
0,299 | ||
4 | 0,373 | 17,6 | 0,096 |
0,297 | ||
5 | 0,433 | 17,5 | 0,129 |
0,298 | ||
6 | 0,581 | 17,4 | 0,172 |
0,296 | ||
7 | 0,778 | 17,4 | 0,230 |
0,296 | ||
8 | 1,040 | 17,2 | 0,311 |
=(17,4-17,0)/2=0,2 | 0,299 | =0,2/1,040=0,19 |
9 | 1,400 | 17,0 | 0,415 |
0,25 | 0,296 | 0,18 |
10 | 1,870 | 16,7 | 0,550 |
0,3 | 0,294 | 0,16 |
11 | 2,500 | 16,4 | 0,740 |
0,4 | 0,296 | 0,16 |
12 | 3,350 | 15,9 | 0,995 |
0,6 | 0,297 | 0,18 |
13 | 4,490 | 15,2 | 1,325 |
0,8 | 0,295 | 0,18 |
14 | 6,00 | 14,3 | 1,755 |
1 | 0,293 | 0,17 |
15 | 8,00 | 13,2 | 2,350 |
1,45 | 0,294 | 0,18 |
16 | 10,70 | 11,4 | 3,050 |
1,95 | 0,285 | 0,18 |
17 | 14,1 | 9,3 | 3,600 |
2,4 | 0,255 | 0,17 |
18 | 17,9 | 6,6 | 2,100 |
1,35 | 0,117 | 0,075 |
19 | 18,3 | 6,5 | 0,200 |
non calculé | 0,011 | non calculé puisque rS |
20 | 18,3 | 6,5 |
Le tableau ci-dessus a été obtenu en adaptant partiellement la présentation de données figurant dans le chapitre fermentation modelling de Sinclair C. G., Cantero D. 1990. In: Fermentation a practical approach. Eds.: McNeil B., Harvey L. M.; IRL PRESS at Oxford University Press, New York, pp. 65-112.
On suppose que la composition élémentaire de la biomasse est CH1,9O0,4N0,28P0,02 plus 4% de la masse sous forme de minéraux.. On a vu alors au paragraphe 1 que le rendement stoechiométrique de conversion de NO3- en biomasse est de 1,5 g de biomasse g-1 de NO3-. Les nitrates sont la source d'élément azote N de la biomasse (N des acides aminés, des bases azotées ...). NO3- n'est pas impliqué dans les processus énergétiques. On peut penser que les pertes en molécules azotées liées au "turnover" physiologique des molécules ne sont pas élevées. Il n'est donc pas ridicule de penser a priori que le rendement in vivo soit très proche du rendement limite stoechiométrique.
On peut tester très simplement l'idée ci-dessus en calculant le rendement global de conversion de NO3- en biomasse sur la durée de 19 h qui a conduit à l'épuisement du glucose. Le calcul donne YX/NO3 global expérimental = (18,3-0,1)/(17,8-6,5) = 1,6 g de biomasse g-1 NO3-. Le résultat expérimental de rendement global est à 1,6 g/g, meilleur que la valeur limite stoechiométrique à 1,5 g/g, ce qui est impossible. Mais on travaille avec des valeurs expérimentales arrondies de données de composition et de données de culture et entachées des erreurs expérimentales et 1,5 # 1,6. Conclusion : le rendement réel est quasi égal au rendement stoechiométrique compte tenu des erreurs expérimentales (sur les mesures de NO3- et de composition élémentaire des cellules). L'hypothèse d'un facteur de maintenance très faible apparaît très pertinente.
On reprend le modèle de base exposé au paragraphe 1.
$$r_{NO3-}=\frac{1}{Y_{X/NO3}^{'}}r_X+m_S[X]$$Si on considère que le terme de maintenance est négligeable en phase de croissance (facteur de maintenance très proche de zéro), ainsi que les considération du §3.2 le laissent penser, le modèle se simplifie en :
$$r_{NO3-}\simeq\frac{1}{Y_{X/NO3}^{'}}r_X$$On peut alors tracer le graphe rS=f(rX) pour tester le modèle. On devrait obtenir une droite linéaire (passant par O(0,0)) si Y’X/NO3 est constant pendant la culture ce qui ne paraît pas être déraisonnable. Et on obtient :
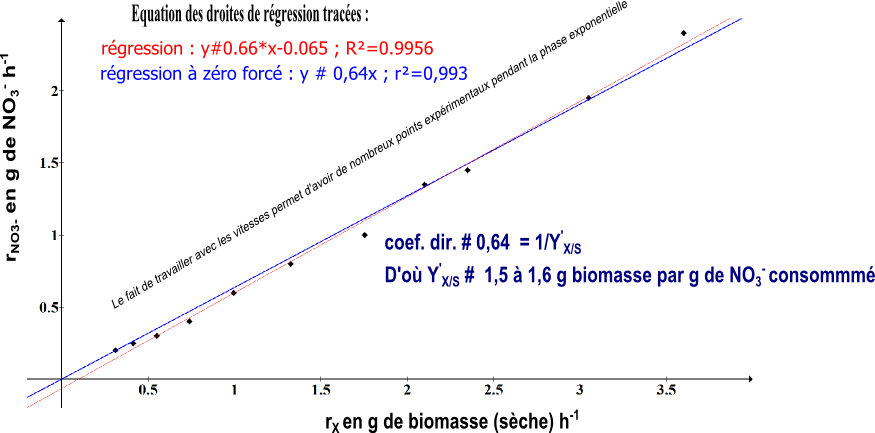
Le modèle physiologique proposé apparaît confirmé. Il s'agit de bien comprendre la signification des résultats obtenus. Y'X/NO3 # 1,5 g de biomasse g-1 de NO3- signifie qu'avec 1 g de NO3-, les cellules peuvent créer 1,5 g de biomasse "neuve". Et le NO3- n'est utilisé que comme source de N, élément constitutif des acides aminés protéines, des bases azotées des nucléotides... Et le facteur de consommation lié à la maintenance est très très faible (puisque considéré comme nul dans le modèle et ça a "collé" avec les données expérimentales).
Et évidemment, compte-tenu de notre modèle, rendement stoechiométrique de conversion de NO3- en biomasse = rendement instantané en chaque temps de la culture en croissance # 1,5 g de biomasse g-1 de NO3- # rendement global de conversion lors de la croissance.
Reprenons le modèle général de consommation pour conversion en biomasse comme exposé au paragraphe 1 :
$$r_{NO3-}=\frac{1}{Y_{X/NO3}^{'}}r_X+m_S[X] $$Il donne mathématiquement (en divisant par [X]) et en dehors de toute considération physiologique :
$$\frac{r_{NO3-}}{[X]}=\frac{1}{Y_{X/NO3}^{'}}{\frac{r_X}{[X]}}+m_S $$On devrait donc trouver que la vitesse spécifique de consommation des NO3- est une fonction affine de la vitesse spécifique de croissance (µ). Et la pente et l'ordonnée à l'origine donneront le coefficient de conversion pour la croissance et le coefficient de maintenance.
Voilà un graphe à essayer qui semble très pertinent ! Malheureusement on n'a que très peu de points expérimentaux à disposition puisqu'on ne dispose que de 2 points expérimentaux en phase de ralentissement et des points en phase exponentielle en théorie tous confondus.
Vous pouvez ouvrir un cadre qui propose un graphe \(\frac{r_{NO3-}}{[X]}=f(\mu)\) réalisé avec les données disponibles et qui montre bien l'insuffisance de données expérimentales pour des vitesses spécifiques de croissance variées. Un étude plus sophistiquée serait nécessaire, chémostats... Etude expérimentale \(\frac{r_{NO3-}}{[X]}=f(\mu)\)".